





![]()
![]()
![]()
BibTeX | RIS | EndNote | Medlars | ProCite | Reference Manager | RefWorks
Send citation to:
URL: http://jams.arakmu.ac.ir/article-1-6202-fa.html



2- گروه زیستشناسی، دانشکده علوم، دانشگاه محقق اردبیلی، اردبیل، ایران. ،
زمینه و هدف
سندروم تخمدان پلیکیستیک (PCOS) یکی از مهمترین عوامل اختلال در تخمکگذاری و ناباروری زنان است که با علائمی نظیر سیکلهای قاعدگی نامنظم (الیگوآمنوره) یا عدم تخمکگذاری (آمنوره)، افزایش سطوح سرمی آندروژنها و افزایش نسبت هورمون لوتئینهکننده (LH) به هورمون محرک فولیکولی (FSH) همراه است [2 ،1].
آدیپونکتین در بافت چربی، هیپوتالاموس، گنادها، عضلات اسکلتیپ و قلب سنتز میشود [4 ،3]. آدیپونکتین اثرات فیزیولوژیکی خود را از طریق اتصال به دو گیرنده AdipoR1 و AdiopoR2 اعمال میکند. گیرندههای آدیپونکتین در عضلات اسکلتی، قلب، کبد، مغز، سلولهای بتا پانکراس، بیضه، سلولهای تکا و گرانولوزای تخمدان بیان میشوند [4 ،3]. نتایج مطالعات پیشین نشان میدهد که سطوح پلاسمایی آدیپونکتین در افراد چاق، بیماران دیابتی و قلبی-عروقی پایینتر از افراد سالم است [6 ،5]. نقص در تولید آدیپونکتین منجر به ایجاد مقاومت به انسولین، عدم تحمل گلوکز و اختلال در متابولیسم لیپیدها میشود [8 ،7]؛ در حالی که آگونیستهای گیرنده آدیپونکتین به عنوان داروی درمانی برای بیماریهای مرتبط با چاقی نظیر دیابت و سندرم متابولیک عمل میکنند [3]. هورمون آدیپونکتین ترشح GnRH/LH را با فعال کردن مسیر پروتئین کیناز فعالشونده cAMP (AMPK) مهار میکند [9] و سطوح سرمی آدیپونکتین در زنان PCOS کمتر از افراد سالم است [10 ،8].
الدوپا پیشساز نوروترانسمیترهای دوپامین، اپینفرین و نوراپینفرین است که از اسیدآمینه L –تیروزین توسط تیروزین هیدروکسیلاز ایجاد میشود. نامهای تجاری آن pharmacopa و madopar است و از نظر کلینیکی در درمان بیماریهای مرتبط با کاهش آزادسازی دوپامین استفاده میشود[11-14]. هنگام تزریق محیطی، دوپامین به علت عدم توانایی عبور از سد خونی-مغزی نمیتواند مستقیماً روی دستگاه عصبی مرکزی اثر بگذارد، درحالیکه پیشسازهای دوپامین (تیروزین یا الدوپا) یا آگونیستهای سنتتیک دوپامین نظیر SKF-38393 توانایی عبور از سد خونی مغزی را دارند [14]. گیرندههای دوپامین روی 50 درصد نورونهای GnRH بیان میشوند. دوپامین و آگونیستهای گیرندههای آن سبب مهار فعالیت هیپوتالاموس- هیپوفیز- گنادها (HPG) و کاهش آزادسازی GnRH/LH میشوند [16 ،15]. همچنین نشان داده شده است که تخمدان انشعابات دوپامینی را دریافت میکند و گیرندههای دوپامین و آنزیم تیروزین هیدروکسیلاز در تخمدان و نواحی مختلف مغز بیان میشود [18 ،17] و استفاده از آنتاگونیستهای گیرندههای دوپامین سبب عدم تخمکگذاری و ایجاد مرحله استروس دائمی در موشهای صحرایی میشود [17]. در افراد PCOS کاهش نسبی در میزان دوپامین مشاهده شده است که میتواند توضیحی برای افزایش ترشح GnRH و درنتیجه افزایش سطح سرمی LH در بیمارانPCOS باشد [19]. در تحقیق حاضر، اثرات تزریق داخل صفاقی الدوپا و آنتاگونیستهای گیرنده دوپامینی شامل SCH23390 هیدروکلراید به عنوان آنتاگونیست گیرنده D1 و سولپرید به عنوان آنتاگونیست گیرنده D2 بر ترشح هورمون LH و میانگین بیان نسبی ژن آدیپونکتین در هیپوتالاموس و تخمدان در موشهای صحرایی ماده مبتلا به PCOS القایی با استرادیول والرات بررسی شد.
مواد و روشها
واحدهای آزمایشی: تخقیق حاضر از نوع تجربی بنیادی است. برای انجام این تحقیق، 20 موش صحرایی ماده نژاد ویستار به وزن 220-180 گرم خریداریشده از مرکز تحقیقات علوم اعصاب دانشگاه شهید بهشتی استفاده شد. در تمامی مدت آزمایش، آب و غذای مخصوص موش صحرایی آزادانه در اختیار حیوانات قرار گرفت. دمای محل نگهداری حیوان در حد 2±22 درجه سانتیگراد بود و حیوانات همواره تحت شرایط آزمایشگاهی 12 ساعت روشنایی و 12 ساعت تاریکی بودند. شروع روشنایی ساعت 7 صبح بود.
بررسی واژیناسیون و القای سندروم تخمدان پلیکیستیک: ابتدا موشها به مدت دو هفته برای خو گرفتن به شرایط آزمایشگاه در قفسها با آب و غذای کافی و دمای مناسب نگهداری شدند؛ سپس سیکل استروس آنها به مدت دو هفته برای بررسی منظم بودن سیکل زیر نظر گرفته شد. پس از مشاهده دو دور سیکل استروس مرتب (بهترتیب پرو استروس، استروس، مت استروس و دی استروس) موشهای صحرایی برای شروع آزمایش آماده شدند. برای القای پلیکیستیک، تزریق عضلانی استرادیول والرات (پودر تهیهشده از شرکت ابوریحان، ایران) با دوز 2 میلیگرم در 0/2 میلیلیتر روغن کنجد (شرکت باریج اسانس، ایران) در مرحله استروس انجام شد. موشها به مدت 60 روز در شرایط آزمایشگاهی با آب و غذای کافی قرار گرفتند. اسمیر واژنی برای بررسی القای PCOS صورت گرفت. از روز 30 تثبیت حالت پلیکیستیک اتفاق افتاد و بر اساس بررسیهای واژیناسیون و مشاهده مرحله استروس پایدار، القای PCOS تأیید شد.
تزریق داروها: برای انجام این آزمایش 15 موش صحرایی PCOS به طور تصادفی به سه گروه 5 تایی تقسیم شدند. گروه اول سالین، گروه دوم 100mg/kg الدوپا (شرکت سیگما، آمریکا) و گروه سوم تزریق همزمان SCH23390 1mg/kg هیدروکلراید (شرکت سیگما، آمریکا)، 10 mg/kg سولپرید و 100mg/kg الدوپا را در حجم 0/5 میلیلیتر از طریق تزریق داخل صفاقی در بازه زمانی 9/30- 9 صبح به مدت دو هفته دریافت کردند. پنج موش صحرایی که در مرحله استروس روغن کنجد را دریافت کرده بودند، بعد از 60 روز به عنوان گروه کنترل منفی، تزریق داخل صفاقی سالین را دریافت کردند. قابل ذکر است که در گروههای دریافتکننده تزریق آنتاگونیست و الدوپا، آنتاگونیستها 10 دقیقه قبل از الدوپا تزریق شد. مقادیر الدوپا و آنتاگونیستها بر اساس تحقیقات پیشین انتخاب شده است [21 ،20].
سنجش نمونههای سرمی: میانگین غلظت سرمی هورمون LH با استفاده روش رادیوایمنواسی (RIA) بر طبق دستورالعمل کیت سنجش هورمون LH ویژه موش صحرایی (Institute of Isotopes Co., Hungary) اندازهگیری شد.
جداسازی نمونههای بافتی: حیوانات با استفاده از کتامین (mg/Kg BW 80) زایلسین (10mg/KgBW) بیهوش شدند. سر حیوانات جدا و جمجمه آن شکافته شد و مغز بلافاصله خارج گردید. سطح شکمی مغز به سمت بالا قرار گرفت و برشی به ضخامت mm 4 حاوی هیپوتالاموس (از جلو از مجاورت اپتیک کیاسما، از پشت تا مجاورت دستگاه پستانی-تالاموسی و به طور جانبی تا شیار هیپوتالاموسی) تهیه شد. هیپوتالاموس و نمونههای تخمدانی راست جداشده بلافاصله در نیتروژن مایع فریز شدند و در دمای 80- تا زمان استخراج RNA نگهداری شدند.
بررسی میزان بیان ژنی با استفاده از روش ریل تایم-PCR: نمونههای هیپوتالاموسی و تخمدان با استفاده از pureZol (Bio Rad Co, U.S.A) و دستگاه هوموژنایزر هوموژن شدند. RNA مطلق نمونهها با استفاده از کلروفرم (Merck Co, Germany)، اپزوپروپانول (Merck Co, Germany) و اتانول 75% طبق دستورالعمل کیت pureZol (Bio Rad Co, U.S.A) استخراج شد. غلظت RNA با استفاده از دستگاه نانودراپ (Themo Scientific, U.S.A) تعیین شد. cDNA تکرشتهای با استفاده یک میکروگرم RNA مطلق، 1 میکرولیتر پرایمر الیگوتیمین (40Mµ)، 1 میکرولیتر مخلوط نوکلئوتید تری فسفاتها (10mMdNTP)، 1 میکرولیتر آنزیم (unit100) M-MuLV به همراه 2 میکرولیتر بافر 10 X M-MuLV بر طبق دستورالعمل کیت سنتز cDNA (Vivantis Co., Malaysia) با استفاده از دستگاه ترمال سایکلر (Bio RAD, U.S.A) سنتز شد. تعیین سطح mRNA گلیسر آلدئید 3 فسفو دهیدروژناز (GAPDH) توسط روش RT- PCR کمی برای نرمال کردن نمونههای mRNA آدیپونکتین استفاده شد.
پس از سنتز cDNA قطعات موردنظر ژنها بر حسب دستورالعمل کیت سایبرگرین ریل تایم پیسیآر شرکت تاکارا (Takara Bio Inc., Japan) و با استفاده از دستگاه ریل تایم پیسیآر روتر ژن مدل 6000 ( Rotor Gene 6000, Corbette, Korea) تکثیر شدند. برنامه زمانی برای واکنش پیسیآر کمی شامل یک چرخه (C °95 برای 2 دقیقه) و 40 چرخه (C °95 برای 5 ثانیه، C °60 برای 25 ثانیه و C °60 برای 20 ثانیه) بود. پرایمرها از شرکت ژن فناوران ایران تهیه شدند. توالیهای الیگونوکلئوتیدی ویژه برای پرایمرهای سنس و آنتیسنس GAPDH و آدیپونکتین بهترتیب برابر با GAPDH sense: 5′- AAGTTCAACGGCACAGTCAAG -3′ and GAPDH antisense: 5′- CATACTCAGCACCAGCATCAC -3′; adiponectin sense: 5′- AATCCTGCCCAGTCATGAAG -3′and adiponectin antisense: 5′- CATCTCCTGGGTCACCCTTA -3′. است. محصولات GAPDH و آدیپونکتین حاصل بهترتیب 120 و 214 جفت باز هستند. دادههای بهدستآمده برای تعیین بیان نسبی ژن آدیپونکتین نسبت به GAPDH با روش دلتا دلتا سی تی طبق فرمول -ΔΔCt 2محاسبه شدند.
تجزیهوتحلیل آماری: دادههای حاصل از فرمول ΔΔCt- 2 با استفاده از نرمافزار SPSS (Version 16) و با استفاده از آزمون آنوای یکطرفه آنالیز شدند. مقایسه میانگین دادهها با استفاده از آزمون تعقیبی توکی انجام شد. نتایج حاصل به صورت میانگین±انحراف معیار میانگین±SEM ارائه شدند. نمودارها با استفاده از نرمافزار اکسل رسم شد. در تمام آنالیزهای آماری نتایج با P≤0/05 معنیدار گزارش شدند.
نتایج
نتایج اثرات تزریق سالین، الدوپا یا تزریق همزمان SCH23390 هیدروکلراید، سولپرید و الدوپا بر میانگین غلظت سرمی LH در گروه کنترل منفی و گروههای PCOS در جدول شماره 1 خلاصه شده است.

میانگین بیان نسبی ژن آدیپونکتین در تخمدان و هیپوتالاموس گروه PCOS نسبت به گروه کنترل منفی کاهش معنیدار نشان داد (0/001=P برای تخمدان، 0/015=P برای هیپوتالاموس) (تصویر شماره 1).
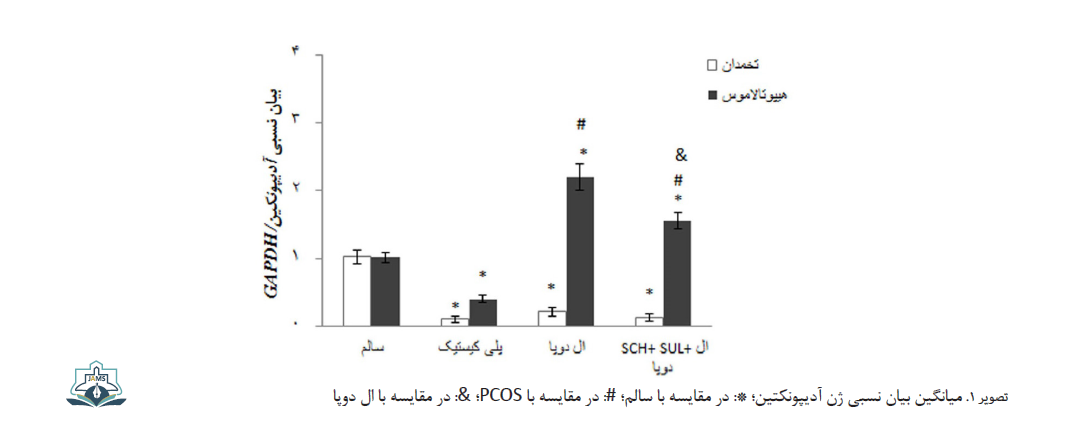
میانگین بیان نسبی ژن آدیپونکتین در تخمدان و هیپوتالاموس گروه PCOS دریافتکننده الدوپا نسبت به گروه PCOS بهترتیب افزایش بیمعنی (0/924=P) و افزایش معنیدار (0/001>P) پیدا کرد (تصویر شماره 1). در گروه تزریق همزمان SCH23390 هیدروکلراید، سولپرید و الدوپا میانگین بیان نسبی ژن آدیپونکتین در تخمدان و هیپوتالاموس در مقایسه با گروه PCOS دریافتکننده الدوپا بهترتیب کاهش بیمعنی (0/948=P) و کاهش معنیدار (0/025=P) نشان داد (تصویر شماره 1).
بحث
نتایج حاصل از این تحقیق نشان داد که در موشهای صحرایی PCOS غلظت سرمی LH در مقایسه با گروه کنترل منفی از نظر آماری به طور معنیداری افزایش یافت. میانگین بیان نسبی ژن آدیپونکتین در تخمدان و هیپوتالاموس موشهای صحرایی PCOS در مقایسه با گروه کنترل منفی از نظر آماری به طور معنیداری کاهش یافت. نتایح حاصل منطبق بر تحقیقات پیشین درباره میزان ترشح آدیپونکتین در وضعیت PCOS در انسان و جوندگان است. تحقیقات پیشین نشان داده است که غلظت سرمی آدیپونکتین در زنان مبتلا به PCOS در مقایسه با افراد سالم کاهش مییابد و زنان چاق PCOS در مقایسه با زنان چاق غیر پلیکیستیک، سطوح سرمی کمتری از آن را دارند [22 ،8]. کاهش سطح آدیپونکتین در زنان PCOS ممکن است در نتیجه افزایش تولید آندروژنها به دلیل کاهش اثرات مهاری آدیپونکتین روی سلولهای تکا باشد [23]؛ زیرا نشان داده شده است که هیپرآندروژنیسم و چاقی در کاهش سطوح پلاسمایی آدیپونکتین و ایجاد مقاومت به انسولین نقش مهمی ایفا میکنند که از مشخصههای اصلی PCOS است [25 ،24].
نتایج تحقیق حاضر نشان داد که تزریق الدوپا سبب افزایش معنیدار میانگین بیان نسبی ژن آدیپونکتین در هیپوتالاموس گروه PCOS در مقایسه با موشهای صحرایی کنترل منفی میشود. تزریق الدوپا میانگین بیان نسبی ژن آدیپونکتین را در تخمدان موشهای صحرایی PCOS در مقایسه با موشهای صحرایی کنترل منفی به طور معنیداری افزایش نداد. نتایج حاصل منطبق بر تنها تحقیق موجود در مورد اثرات دوپامین بر ترشح آدیپونکتین است که در آن محققان اثرات دوپامین بر سلولهای آدیپوسیت انکوبهشده در محیط کشت در شرایط in vitro را بررسی و درنتیجه اثرات تحریکی دوپامین بر ترشح آدیپونکتین از این سلولها را گزارش کردند [22]. همچنین نتایج حاضر نشان داد که تزریق همزمان هر دو نوع آنتاگونیست شامل SCH23390 هیدروکلراید و سولپرید به بلوکه کردن اثرات تحریکی الدوپا بر میانگین بیان نسبی ژن آدیپونکتین در هیپوتالاموس موشهای صحرایی مبتلا به PCOS منجر میشود؛ درحالیکه تزریق همزمان هر دو نوع آنتاگونیست میانگین بیان نسبی ژن آدیپونکتین در تخمدان را تحت تأثیر قرار نداد.
در مورد نقش مسیر دوپامینرژیکی بر فعالیت مسیر عصبی آدیپونکتین اطلاعات زیادی در دسترس نیست و یافتن دقیق این مسیرها نیاز به تحقیقات آتی بیشتری دارد، ولی میتوان بر اساس تحقیقات پیشین و در نظر گرفتن فاکتورهای اصلی دخیل در پاتوژنز PCOS نظیر هیپرآندوژنیسم، مقاومت به انسولین، افزایش ترشح پرولاکتین و غیره احتمال داد که مسیرهای عصبی واسطهای مختلفی ممکن است در اعمال اثرات تحریکی الدوپا بر بیان ژن آدیپونکتین در هیپوتالاموس نقش داشته باشد [31-23]. پرولاکتین با مهار ترشح GnRH موجب مهار ترشح پالسی گنادوتروپینها میشود [26]. پرولاکتین همچنین روند سنتز آندروژنهای آدرنال را تحریک میکند. افزایش آندروژنها میتواند سبب افزایش وزن و پرمویی شود [26]. دوپامین کنترلکننده مهاری پرولاکتین است و تزریق دوپامین موجب سرکوب پرولاکتین در مردان و زنان سالم و همچنین بیماران هیپرپرولاکتینمیا میشود [22].
از طرفی نشان داده شده است که بین کاهش سطوح بیان آدیپونکتین و ایجاد مقاومت به انسولین ارتباط مستقیمی وجود دارد [8 ،7] و افزایش پرولاکتین منجر به ایجاد مقاومت به انسولین میشود [27]؛ همچنین نیلسون و همکاران در سال 2005 نشان دادند که پرولاکتین ترشح آدیپونکتین را در بافت چربی انسان در شرایط in vitro و در جوندگان در شرایط in vivo بهطور قابلتوجهی مهار میکند [26]. با در نظر گرفتن یافتههای این تحقیقات میتوان فرض کردکه کاهش سنتز پرولاکتین با تزریق الدوپا ممکن است یکی از مکانیسمهای احتمالی در ایجاد اثرات تحریکی دوپامین بر بیان ژن آدیپونکتین در این تحقیق باشد.
با در نظر گرفتن اثر مهاری دوپامین بر آندروژنهایی مثل تستوسترون و استرادیول و با توجه به تحقیقات در زمینه تأثیر آندروژنها بر غلظت آدیپونکتین سرم مشخص شده است که تزریق تستوسترون به مردان به طور مستقیم تولید آدیپونکتین را از بافت آدیپوسیت سرکوب میکند و نتایج نشان میدهد که آندروژنها باعث کاهش آدیپونکتین پلاسما میشود و هیپوآدیپونکتینمیا ممکن است با خطرات احتمالی مقاومت به انسولین در مردان مرتبط باشد [29 ،28]. پس کاهش آندروژنها از طریق تزریق دوپامین میتواند یکی دیگر از مکانیسمهای احتمالی در ایجاد اثرات تحریکی دوپامین بر آدیپونکتین باشد. سطوح آدیپونکتین سرم به عنوان یک سیتوکین ضدالتهابی و ضددیابتی دارای ارتباط نزدیک با مقاومت به انسولین است و مطالعات از یک ارتباط معکوس بین آدیپونکتین و مقاومت به انسولین بحث میکنند.
با توجه به اینکه مقاومت به انسولین یکی از مؤلفههای اصلی PCOS محسوب میشود، به نظر میرسد بررسی ارتباط آدیپونکتین و انسولین کمک شایانی به چگونگی اثرات تحریکی دوپامین بر ترشح آدیپونکتین در مدلهای پلیکیستیک کند. انسولین به عنوان مهارکننده سطوح آدیپونکتین در انسانها و حیوانات و تنظیمکننده ترشح آدیپونکتین شناخته شده است [30]؛ همچنین نوعی ارتباط خطی منفی و معنیدار بین سطوح آدیپونکتین و گلوکز ناشتا مشاهده شده است. در این زمینه برگ و همکاران با استناد به یافتههای خود اظهار داشتند که افزایش آنی در سطوح پلاسمایی آدیپونکتین موجب کاهش سطوح گلوکز پلاسما به واسطه مهار بیان آنزیمهای گلوکونئوژنز کبدی در موشهای دیابتی میشود [31]؛ همچنین مطالعات پیشین اثرات مهاری انسولین بر آدیپونکتین را مطرح کردهاند [30]؛ بنابراین کاهش ترشح انسولین توسط دوپامین را میتوان مسیر واسطه مهم دیگری برای اثرات تحریکی مسیر دوپامینرژیکی بر بیان ژن آدیپونکتین در نظر گرفت. باوجوداین، برای یافتن نقش دقیقتر مسیر دوپامینرژِیکی در کنترل ترشح و بیان ژن آدیپونکتین پیشنهاد میشود که اثرات تزریق بطن سوم مغزی یا تزریق داخل هسته قوسی (ARC) یا ناحیه پریاپتیک میانی دوپامین، آگونیستها و آنتاگونیستهای مختلف گیرنده دوپامین بر میانگین غلظت سرمی و بیان ژن آدیپونکتین در هیپوتالاموس و بافت چربی یا اثرات دوپامین بر نوروپپتیدهای کنترلکننده سنتز آدیپونکتین نظیر گرلین، کیسپپتین و غیره در موشهای صحرایی PCOS بررسی شود.
نتیجهگیری
القای PCOS در موشهای صحرایی سبب افزایش غلظت سرمی LH و کاهش بیان ژن آدیپونکتین در تخمدان و هیپوتالاموس شد. الدوپا اثرات مهاری بر ترشح LH و اثرات تحریکی بر بیان ژن آدیپونکتین در هیپوتالاموس موشهای صحرایی PCOS اعمال کرد. الدوپا اثرات تحریکی بر بیان ژن آدیپونکتین در تخمدان موشهای صحرایی PCOS نداشت. آنتاگونیستهای گیرنده دوپامینی شامل SCH23390 هیدروکلراید و سولپرید اثرات مهاری الدوپا بر ترشح LH و اثرات تحریکی آن بر بیان ژن آدیپونکتین در هیپوتالاموس موشهای صحرایی PCOS را بلوکه کرد. احتمال دارد که افزایش فعالیت نورونهای دوپامینرژیکی در کنترل اختلالات اندوکرینی ناشی از کاهش ترشح آدیپونکتین در بیماران PCOS مؤثر واقع شود.
ملاحظات اخلاقی
پیروی از اصول اخلاق پژوهش
این تحقیق را کمیته پژوهشی دانشگاه محقق اردبیلی (کد: 1 125 - 95) تأیید کرده است.
حامی مالی
نتایج این تحقیق مستخرج از پایاننامه دانشجوی کارشناسی ارشد رشته فیزیولوژی جانوری سرکار خانم خدیجه حقیقت گللو است. نویسندگان از حمایتهای مالی و معنوی معاونت پژوهش و فناوری دانشگاه محقق اردبیلی در انجام این پژوهش سپاسگزاری میکنند.
مشارکت نویسندگان
تمــام نویســندگان در آمادهســازی ایــن مقالــه مشــارکت داشــتهاند.
تعارض منافع
نویسندگان اعلام میکنند که هیچگونه تضاد منافعی در این پژوهش وجود ندارد.
تشکر و قدردانی
نویسندگان از حمایت های مالی و معنوی معاونت پژوهش و فناوری دانشگاه محقق اردبیلی در انجام این پژوهش سپاسگزاری میکنند. همچنین نویسندگان از جناب آقای دکتر همایون خزعلی از دانشگاه شهید بهشتی برای تامین دستگاهها صمیمانه تشکر و قدردانی میکنند.
References
1.Polak K, Czyzyk A, Simoncini T, Meczekalski B. New markers of insulin resistance in polycystic ovary syndrome. J Endocrinol Invest. 2017; 40(1):1-8. [DOI:10.1007/s40618-016-0523-8] [PMID] [PMCID]
2.Adgi Z, Talaei A, Mohamadi Kelishadi M. [The evaluation of the relationship between hirsutism and insulin resistance in patients with PCOS and idiopathic hirsutism (Persian)]. J Arak Univ Med Sci. 2011; 14(2):51-7. http://jams.arakmu.ac.ir/article-1-953-en.html
3.Dobrzyn K, Smolinska N, Kiezun M, Szeszko k, Rytelewska E, Kisielewska K, et al. Adiponectin: A new regulator of female reproductive system. Int J Endocrinol. 2018; 2018:7965071. [DOI:10.1155/2018/7965071] [PMID] [PMCID]
4.Davoodi B, Zilaei Bouri Sh, Ahangarpor A, Zilaei Bouri M. [Effects of two different physical exercises on plasma levels of adiponectin and resistin in obese and overweight young girls (Persian)]. J Arak Univ Med Sci. 2014; 17(4):27-37. http://jams.arakmu.ac.ir/article-1-2206-en.html
5.Rodriguez-Pacheco F, Martinez-Fuentes AJ, Tovar S, Pinilla L, Tena-Sempere M, Dieguez C, et al. Regulation of pituitary cell function by adiponectin. Endocrinology. 2007; 148(1):401-10. [DOI:10.1210/en.2006-1019] [PMID]
6.Lee B, Shao J. Adiponectin and energy homeostasis. Rev Endocr Metab Disord. 2014; 15(2):149-56. [DOI:10.1007/s11154-013-9283-3] [PMID] [PMCID]
7.Groth SW. Adiponectin and polycystic ovary syndrome. Biol Res Nurs. 2010; 12(1):62-72. [DOI:10.1177/1099800410371824] [PMID] [PMCID]
8.Michalakis KG, Segars JH. The role of adiponectin in reproduction: From polycystic ovary syndrome to assisted reproduction. Fertil Steril. 2010; 94(6):1949-57. [DOI:10.1016/j.fertnstert.2010.05.010] [PMID] [PMCID]
9.Cheng XB, Wen JP, Yang J, Yang Y, Ning G, Li XY. GnRH secretion is inhibited by adiponectin through activation of AMP-activated protein kinase and extracellular signal-regulated kinase. Endocrine. 2011; 39(1):6-12. [DOI:10.1007/s12020-010-9375-8] [PMID]
10.Parillo F, Maranesi M, Mignini F, Marinelli L, Di Stefano A, Boiti C, et al. Evidence for a dopamine intrinsic direct role in the regulation of the ovary reproductive function: In vitro study on rabbit corpora lutea. PLoS One. 2014; 9(8):e104797. [DOI:10.1371/journal.pone.0104797] [PMID] [PMCID]
11.Björklund A, Dunnett SB. Dopamine neuron systems in the brain: An update. Trends Neurosci. 2007; 30(5):194-202. [DOI:10.1016/j.tins.2007.03.006] [PMID]
12.Vallone D, Picetti R, Borrelli E. Structure and function of dopamine receptors. Neurosci Biobehav Rev. 2000; 24(1):125-32. [DOI:10.1016/S0149-7634(99)00063-9]
13.Fontaine R, Affaticati P, Yamamoto K, Jolly C, Bureau C, Baloche S, et al. Dopamine inhibits reproduction in female zebrafish (Danio rerio) via three pituitary D2 receptor subtypes. Endocrinology. 2013; 154(2):807-18. [DOI:10.1210/en.2012-1759] [PMID]
14.Zarabian M, Salehipour F, Ostad SN. The study of dose-response mitogenic effect of L-dopa on the human periodontal ligament fibroblasts cell. Acta Med Iran. 2004; 42(5):363-6. https://acta.tums.ac.ir/index.php/acta/article/view/2752
15.Liu X, Herbison AE. Dopamine regulation of gonadotropin-releasing hormone neuron excitability in male and female mice. Endocrinology. 2013; 154(1):340-50. [DOI:10.1210/en.2012-1602] [PMID]
16.Venegas-Meneses B, Padilla JF, Juárez CE, Morán JL, Morán C, Rosas-Murrieta NH, et al. Effects of ovarian dopaminergic receptors on ovulation. Endocrine. 2015; 50(3):783-96. [DOI:10.1007/s12020-015-0636-4] [PMID]
17.Chaudhari N, Dawalbhakta M, Nampoothiri L. GnRH dysregulation in Polycystic Ovarian Syndrome (PCOS) is a manifestation of an altered neurotransmitter profile. Reprod Biol Endocrinol. 2018; 16(1):37. [DOI:10.1186/s12958-018-0354-x] [PMID] [PMCID]
18.Ayano G. Dopamine: Receptors, functions, synthesis, pathways, locations and mental disorders: Review of literatures. J Ment Disord Treat. 2016; 2(2):1000120. [DOI:10.4172/2471-271X.1000120]
19.Gómez R, Ferrero H, Delgado-Rosas F, Gaytan M, Morales C, Zimmermann RC, et al. Evidences for the existence of a low dopaminergic tone in polycystic ovarian syndrome: Implications for OHSS development and treatment. J Clin Endocrinol Metab. 2011; 96(8):2484-92. [DOI:10.1210/jc.2011-0075] [PMID]
20.Andersson K, Fuxe K, Eneroth P, Härfstrand A, Agnati LF. Involvement of D1 dopamine receptors in the nicotine-induced neuro-endocrine effects and depletion of diencephalic catecholamine stores in the male rat. Neuroendocrinology. 1988; 48(2):188-200. [DOI:10.1159/000125007] [PMID]
21.Grierson JP, James MD, Pearson JR, Wilson CA. The effect of selective D1 and D2 dopaminergic agents on sexual receptivity in the female rat. Neuropharmacology. 1988; 27(2):181-9. [DOI:10.1016/0028-3908(88)90169-4]
22.Borcherding DC, Hugo ER, Idelman G, De Silva A, Richtand NW, Loftus J, et al. Dopamine receptors in human adipocytes: Expression and functions. PloS One. 2011; 6(9):e25537. [DOI:10.1371/journal.pone.0025537] [PMID] [PMCID]
23.Lagaly DV, Aad PY, Grado-Ahuir JA, Hulsey LB, Spicer LJ. Role of adiponectin in regulating ovarian theca and granulosa cell function. Mol Cell Endocrinol. 2008; 284(1-2):38-45. [DOI:10.1016/j.mce.2008.01.007] [PMID]
24.Escobar-Morreale HF, Villuendas G, Botella-Carretero JI, Álvarez-Blasco F, Sanchón R, Luque-Ramírez M, et al. Adiponectin and resistin in PCOS: A clinical, biochemical and molecular genetic study. Hum Reprod. 2006; 21(9):2257-65. [DOI:10.1093/humrep/del146] [PMID]
25.Nishizawa H, Shimomura I, Kishida K, Maeda N, Kuriyama H, Nagaretani H, et al. Androgens decrease plasma adiponectin, an insulin-sensitizing adipocyte-derived protein. Diabetes. 2002; 51(9):2734-41. [DOI:10.2337/diabetes.51.9.2734] [PMID]
26.Nilsson L, Binart N, Bohlooly-Y M, Bramnert M, Egecioglu E, Kindblom J, et al. Prolactin and growth hormone regulate adiponectin secretion and receptor expression in adipose tissue. Biochem Biophys Res Commun. 2005; 331(4):1120-6. [DOI:10.1016/j.bbrc.2005.04.026] [PMID]
27.Daimon M, Kamba A, Murakami H, Mizushiri S, Osonoi Sh, Yamaichi M, et al. Association between serum prolactin levels and insulin resistance in non-diabetic men. PLoS One. 2017; 12(4):e0175204. [DOI:10.1371/journal.pone.0175204] [PMID] [PMCID]
28.Xu A, Chan KW, Hoo RLC, Wang Y, Tan KCB, Zhang J, et al. Testosterone selectively reduces the high molecular weight form of adiponectin by inhibiting its secretion from adipocytes. J Biol Chem. 2005; 280(18):18073-80. [DOI:10.1074/jbc.M414231200] [PMID]
29.Page ST, Herbst KL, Amory JK, Coviello AD, Anawalt BD, Matsumoto AM, et al. Testosterone administration suppresses adiponectin levels in men. J Androl. 2005; 26(1):85-92. [DOI:10.1002/j.1939-4640.2005.tb02876.x]
30.Giahi L, Djazayery A, Rahimy A, Rahmany M, Larijani B. Serum level of adiponectin and its association with insulin sensitivity in overweight diabetic and non-diabetic Iranian men. Iran J Public Health. 2008; 37(2):88-92. https://ijph.tums.ac.ir/index.php/ijph/article/view/2060
31.Berg AH, Combs TP, Du X, Brownlee M, Scherer PE. The adipocyte-secreted protein Acrp30 enhances hepatic insulin action. Nat Med. 2001; 7(8):947-53. [DOI:10.1038/90992] [PMID]
| بازنشر اطلاعات | |
 |
این مقاله تحت شرایط Creative Commons Attribution-NonCommercial 4.0 International License قابل بازنشر است. |
